Structural inheritance: The parent as a developmental template
23 July 2018
Structural inheritance is an often-neglected form of nongenetic inheritance. In their new book, Extended Heredity: A New Understanding of Inheritance and Evolution, authors Russell Bonduriansky and Troy Day describe numerous examples of structural inheritance, where structural features – and their mutilations – are inherited in subsequent generations through self-templating and related processes.
Parental traits scaffold offspring development in many ways and at different levels. For example, at the whole organism level, parental traits provide an ambient environment (and, in some animals, also a cultural context) for development. Scaffolding also occurs at the cellular and molecular level, through inheritance of epigenetic and cytoplasmic factors, and the template-like processes of cell division that shape daughter cell features. This scaffolding is inescapable because all cells come from pre-existing cells (all the way back to the origin of life), and cell components cannot spontaneously assemble themselves to form a new cell. For some cell features that are more or less directly shaped by pre-existing features, we can go beyond a general scaffold for development and talk about structural inheritance.
There are numerous fascinating examples of structural inheritance occurring in protists (single-celled eukaryotes). From the mid-1900’s researchers have been manually disrupting structural elements of these tiny yet complex organisms with extremely interesting results. The protist Difflugia corona has a ring of ‘teeth’ around its circular ‘mouth’. When some teeth are knocked out using a fine glass needle, daughter cells inherit the altered number and shape of teeth for many generations producing new lines with these new teeth patterns.1 Another example is the pattern of cilia on Paramecium aurelia. This protist can reproduce sexually via conjugation where two individuals join temporarily to exchange genetic material. When part of one individual is removed, the other incorporates the remainder of its partner into itself, which changes the pattern of cilia on its surface. This altered pattern can be passed on for hundreds of generations and influences the swimming patterns of the descendent cells.2
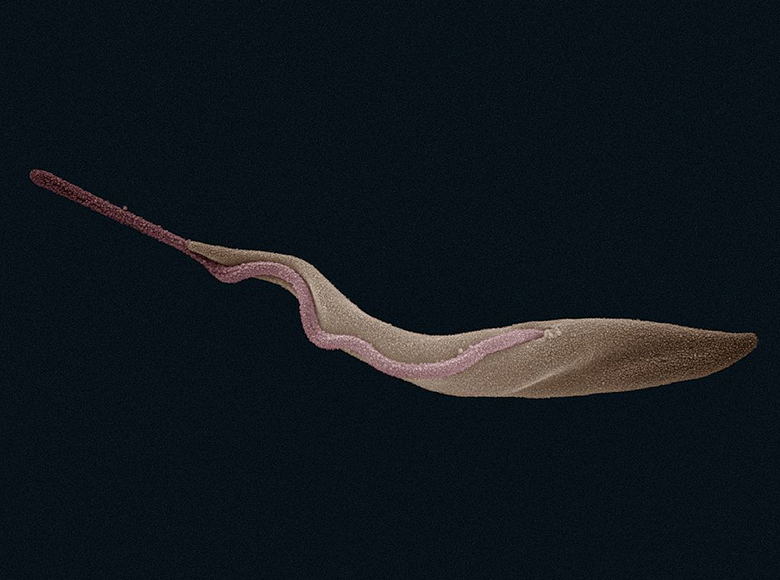
Trypanosoma brucei is the protozoan parasite that causes sleeping sickness in humans. Each cell has a single attached flagellum that runs the length of the cell in a helical pattern. As the new flagellum grows along the daughter cell during cell division, it remains tethered to the parental flagellum, which directs and maintains its path along the daughter cell. This in turn arranges intracellular components and determines cell polarity.3 The cell wall of the diatom Cyclotella meneghiniana provides yet another example of parental templating. These cells have specialized cell wall structures called siliceous valves, one at each pole. During cell division, each daughter cell receives one parental valve from which a new value is formed, faithfully replicating parental valve structure.4
{kind=link}
The mechanisms of structural inheritance go beyond simple templated copying. Take hypotrich ciliates for example. When they are starved, they become cysts – dormant, round, apparently featureless balls. When conditions improve, they emerge from the cysts and take on their previous active form. If the ciliates are experimentally manipulated before encystment, they emerge from the cyst regenerating some elements of the manipulated form.5
The examples above suggest that structural inheritance is pervasive over eukaryotic lineages and that the cortex (surface membrane and underlying proteins) and cytoskeleton of the parent cell shape features of the daughter cell. These and other examples demonstrate striking changes in phenotype without change to genetic sequence, with these phenotypes sometimes inherited over many, many generations. Some important questions arise: What about structural inheritance in multi-celled eukaryotes? Is it involved in inheritance of non-cortical structures? And what are the evolutionary consequences of structural inheritance?
Processes similar to those in single-cell eukaryote cell division probably occur in cell lineages during development of multi-celled organisms. But we don’t yet know if structural variations persist across generations in multi-celled eukaryotes and if they play a role in heredity. There are, however, some suggestions that this might be the case. The structure of the egg cell – both the cortex and the intracellular components – is important for proper embryo development. When egg cell contents are mixed up, embryos develop abnormally. As the egg is made by the mother, it can reflect influences of both the maternal genome and environment, and it’s possible that some of these influences can be passed on to the next generation via egg structure.
Other potential instances of structural inheritance in multi-celled eukaryotes may be found in lipid membranes and prion-like proteins. Cellular membranes are always formed from pre-existing membranes through the insertion of proteins and lipids. Membrane heredity may allow the inheritance of membrane polarity, topology and composition from previous generations.6 Prions are proteins that direct certain normal proteins to fold abnormally, leading to neurodegenerative diseases such as mad cow disease (bovine spongiform encephalopathy). By passing on their abnormal protein folding, prions replicate and disease progresses. It’s possible that there are many normal cytoplasmic proteins with prion-like properties, capable of altering the three-dimensional shape of other proteins. These may be responsive to the environment and inherited through the germline via the gamete cytoplasm.7
There is still a lot of work to be done to fully appreciate the generality and importance of structural inheritance in evolution. We know that it appears to be pervasive across single-cell eukaryotes but it is still unclear whether it occurs trans-generationally in multi-celled organisms. If it does, understanding the evolutionary consequences of this form of extra-genetic inheritance will become an exciting research direction.
Russell Bonduriansky and Troy Day’s extended heredity perspective differs from that of the extended evolutionary synthesis yet it is aligned in many ways. Stay tuned for an upcoming post where we interview them to understand the differences between these perspectives.
1. Jennings HS. 1937. J Exp Zool 77:287-336. 2. Beisson J & Sonneborn TM. 1965. PNAS 53:275-82. 3. Moreira-Leite FF, Sherwin T, Kohl L, Gull K. 2001. Science 294:610-612. 4. Shirokawa Y & Shimada M. 2016. Proc R Soc B 283:20161632. 5. Grimes GW. 1982. Am Zool 22:35-46. 6. Cavalier-Smith T. 2004. CRC Press. 7. Halfmann R & Lindquist S. 2010. Science 330:629-32.
Buy the book
Extended Heredity: A New Understanding of Inheritance and Evolution by Russell Bonduriansky & Troy Day (Princeton University Press, 2018)