Organisms can respond plastically to novel conditions to generate functional variation. The specific adaptive phenotypes generated need not be the direct targets of past selection. Rather, they are often the expression of the more general ability of developmental processes to accommodate novel inputs adaptively, enabling functionally integrated responses to a broad range of conditions (West-Eberhard 2003; Kirschner & Gerhart 2007).
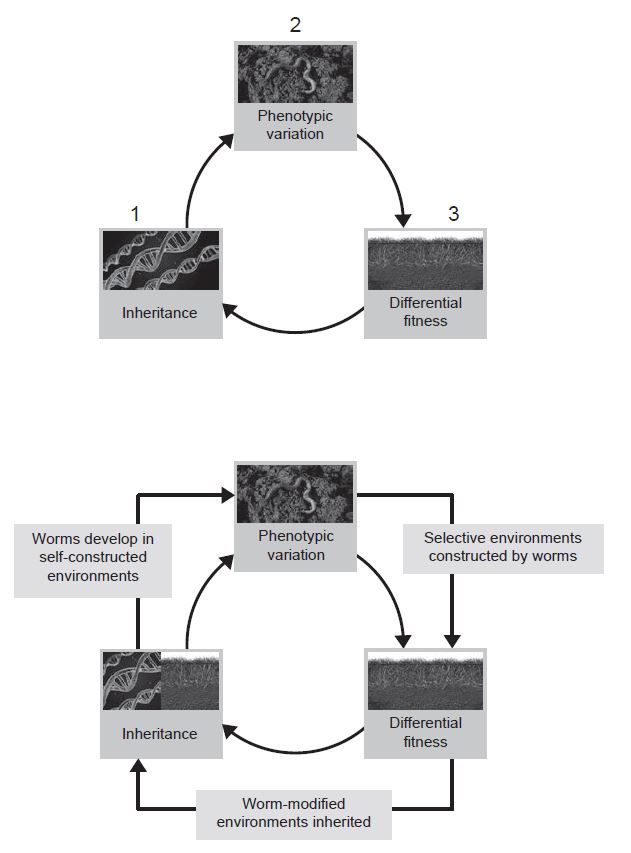
Moreover, through niche construction, environments can be changed by organisms to suit themselves. Consider the example of the earthworms which, despite living on land for millions of years, have retained the characteristic physiology of the freshwater animals from which they evolved (Turner 2000). For instance, earthworms produce large quantities of dilute urine, a trait that on land leaves them highly vulnerable to desiccation. Earthworms can only survive because they process the soil in ways that allow them to draw water into their bodies more effectively. They construct a simulated aquatic environment on land.
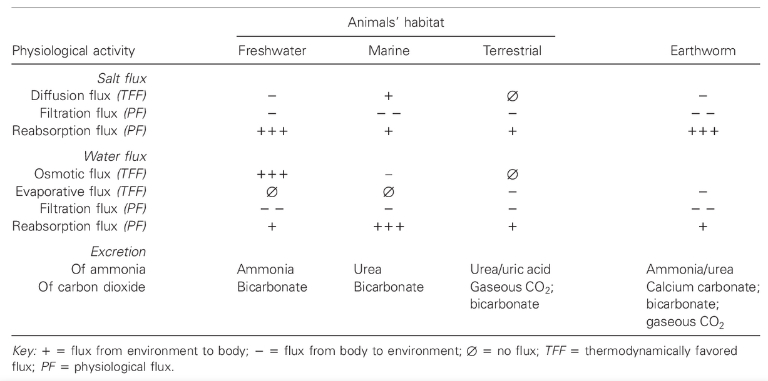
The key point is that the adaptive complementarity of earthworms and soils results to a large extent from the worms changing the soil through niche construction, rather than natural selection changing the worms to a typical terrestrial physiology.
Traditionally, the latter route has not been recognized as an alternative to adaptation – after all, the earthworm’s niche-constructing traits are adaptations too. However, the conventional stance assumes a particular model of biological causation, which advocates of the niche-construction perspective reject (Laland et al. 2011).
From the niche-construction perspective, the current selective environment was itself brought about through earlier niche construction, which caused the bout of selection. The attribution of all causal significance to selection fails to capture the true reciprocal nature of causation in this system.
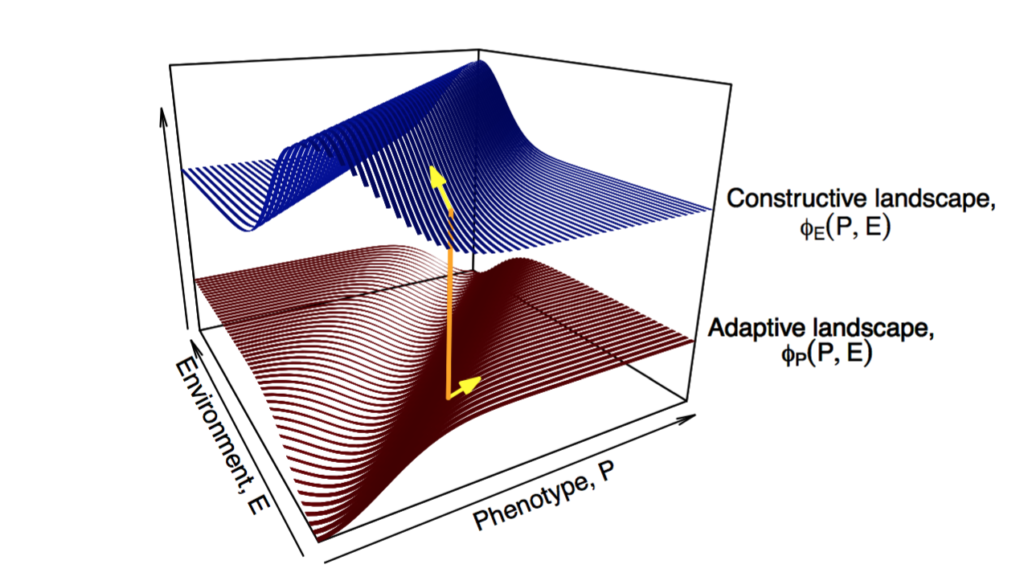
Central to these debates are different assumptions concerning the independence or interdependence of the causes of phenotypic variation, differential fitness and inheritance, which are Lewontin’s (1970) three conditions for evolution by natural selection (Walsh 2015; Uller & Helanterä 2019). Traditionally, evolutionary biologists have assumed these processes are quasi-independent, but in practice they are often causally intertwined.

Harvard evolutionary biologist and Modern Synthesis architect Ernst Mayr (1961) wrote about biological causation, distinguishing between proximate and ultimate causes. Mayr equated proximate causation with immediate factors (e.g. physiology) and ultimate causation with evolutionary explanations (e.g. natural selection). He argued that proximate and ultimate causes addressed different questions and were not alternatives, which remains a valid and non-contentious point.
 Mayr’s account of causation remains widely accepted today, with both positive and negative ramifications. One negative consequence was that Mayr insisted on a strict separation of proximate and ultimate causes:
Mayr’s account of causation remains widely accepted today, with both positive and negative ramifications. One negative consequence was that Mayr insisted on a strict separation of proximate and ultimate causes:
“The clarification of the biochemical mechanism by which the genetic program is translated into the phenotype tells us absolutely nothing about the steps by which natural selection has built up the particular genetic program” (Mayr 1980).
This position was based upon a specific view of development:
“All of the directions, controls and constraints of the developmental machinery are laid down in the blueprint of the DNA genotype as instructions or potentialities” (Mayr 1984).
Mayr and his followers argued that developmental (i.e. ontogenetic) processes were irrelevant to the study of evolution. Niche construction is an ontogenetic process, and advocates of the niche-construction perspective, alongside others, have taken issue with Mayr’s treatment of causation (Laland et al. 2011).
For Mayr, developmental processes could not be regarded as independent causes of evolutionary events, since he believed their properties (which include how organisms construct their niches) were themselves fully explained by prior natural selection. From Mayr’s standpoint, if developmental processes direct evolutionary events, this is only the proximate manifestation of the ultimate cause of natural selection; aspects of development not shaped by selection were assumed to play no evolutionary role.
This stance has made it more difficult for evolutionary biologists to recognize niche construction as an evolutionary process. Niche construction is perceived to have no independent evolutionary significance because, to the extent that it is evolutionarily consequential, it is regarded as fully explained by a preceding cause, natural selection (e.g. Dawkins 2004). Niche-construction effects are treated as ‘extended phenotypes’, whose sole evolutionary role is to affect the probability that gene variants underlying niche construction are passed on to the next generation. Similar reasoning underlies the treatment of niche construction as an indirect genetic effect, with, for example, a mother’s genes affecting her offspring’s phenotype by modifying its environment (Wolf et al. 1998).
There are two major problems with this line of reasoning:
First, as described above, the processes underlying variation, fitness and inheritance are causally intertwined. All organisms have always engaged in niche construction, going right back to the beginning of life. Hence there is no reason to assume that natural selection must be the first cause of any evolutionary episode. Causation in biological systems is reciprocal rather than linear, with natural selection and niche construction codirecting each other.
Second, not all evolutionarily consequential aspects of niche construction (nor all aspects of development, in general) are under genetic control. Niche-construction theorists, like many developmental biologists, regard organisms (and their environmental modification) as under-determined by genes. The changes that organisms bring about do not flow only from their adaptations, but also derive from plasticity, byproducts, and acquired characters.

Evolutionary theory is often viewed as a fusion of Darwinian and Mendelian insights. Notwithstanding the immense progress that these achievements engendered, Richard Lewontin (1983) argues that the legacies of Darwin and Mendel include misleading metaphors that:
are responsible for certain difficulties in biology, difficulties that prevent us from some kinds of further progress and which keep us locked into a rigid framework of thought about the development and evolution of organisms.
Mendel’s view of organisms as the manifestation of autonomous internal ‘factors’ (i.e. genes) with their own laws, germinated into a post-Synthesis metaphor in which ontogeny ‘is seen as an unfolding of a form, already latent in the genes, requiring only an original triggering at fertilization and an environment adequate to allow ‘normal’ development to continue’. We now know that developmental processes are hugely environmentally contingent.
Darwin’s view of organisms as passive objects moulded by the external force of natural selection encouraged a conception of evolution in which ‘the environment “poses the problem”; the organisms posit “solutions”, of which the best is finally “chosen”. The metaphor of selection, inspired by the efficacy of artificial selection, continues to encourage a view of organisms as passive objects upon which external forces act.
Lewontin argues forcefully that these metaphors are misleading. Similar points have been made by Oyama (1985), Griffiths and Gray (2001, 2004), Sultan (2015) and many others.