Niche construction theory contrasts with conventional conceptualizations of evolution in recognizing niche construction as an evolutionary cause or process. The niche-construction perspective seeks to explain the adaptive complementarity of organism and environment in terms of a dynamic, reciprocal interaction between the processes of natural selection and niche construction.

The distinctive aspects of the niche-construction perspective are illustrated by the familiar example of the beaver. Generally, the evolutionary consequences of beaver dam building are modeled in terms of fitness ‘payoffs’ to the underlying genes; selection favors dam-building alleles over their alternatives. According to Richard Dawkins (1982), who exemplifies the traditional standpoint, dams can be viewed as ‘extended phenotypes’Biological adaptations expressed outside of the body of the organism. – adaptations expressed outside of the constructor’s body, but that evolve in a manner no different from other adaptations.
From the niche-construction perspective this stance is unsatisfactory, both because it misses part of the causal story and because it discourages consideration of additional forms of selective feedback. When a beaver builds a dam, creating a lake and influencing river flow, it not only affects the propagation of dam-building genes but dramatically changes its local environment, affecting nutrient cycling, decomposition dynamics, the structure of the riparian zone, and plant and community composition and diversity. It follows that beaver dam building must also transform selection acting on a host of other beaver traits. The active agencyThe capacity of living organisms to act on, and in, their world, and to modify their experience of it, including in ways that are neither predetermined, nor random. of beavers in constructing these modified selection pressures and thereby acting as co-directors of their own evolution (not to mention that of other species) currently goes unrecognized.
Often the modifications produced by niche-constructing organisms persist for longer than the individual constructors, continuing to modulate the impact of environment on subsequent generations, a legacy described as an ‘ecological inheritance’The inheritance, via an external environment, of one or more natural selection pressures previously modified by niche-constructing organisms.. Descendants of the dam builder will experience modified selection so long as the dam, lake, and transformed environment remain. Given that dams are frequently maintained by families of beavers for decades, that could be considerably longer than the lifetime of an individual beaver. In addition to genes, offspring inherit a modified selective environment.
While it is possible to study this feedback using established evolutionary theory, for instance through models of habitat selection, co-evolution or maternal effects, nonetheless the magnitude and significance of niche construction remains underappreciated, in part because standard evolutionary theory does not encourage attention to such phenomena.
Conversely by treating niche construction as an evolutionary process – one that is influenced by, but not solely reducible to, earlier natural selection – niche construction theory generates novel hypotheses and new methods that stimulate research. The perspective is rapidly growing, precisely because researchers from multiple disciplines find it useful.
Several factors render niche construction of evolutionary significance:
(1) Niche construction biases natural selection
Organisms do not modify environments at random. Rather, they create environmental states, such as nests, burrows or benign conditions, that are adaptive for the constructor or its descendants. They can also destroy environments, or produce conditions that impact negatively on fitness. Studies show that environmental features constructed by organisms have different properties, and generate different patterns of selection, to aspects of environments that change independently of the organism (Clark et al. 2020). For instance, selection in response to environmental components regulated by organisms is consistently weaker and less variable than selection arising from autonomous aspects of environments. Organisms modify environments in distinctive, nonrandom ways, thereby imposing a systematic bias on natural selection.
The orderly nature of niche construction follows largely because it has evolved through earlier natural selection. However, this does not mean it can be disregarded. Organisms are influenced, but not determined, by their genes, and their activities are shaped by developmental information-gaining processes as well as natural selection. Organisms are not merely objects through which the causal explanatory power of natural selection flows; rather they are active agents that transduce and filter genetic inputs that derive from prior selection, as well as environmental inputs.
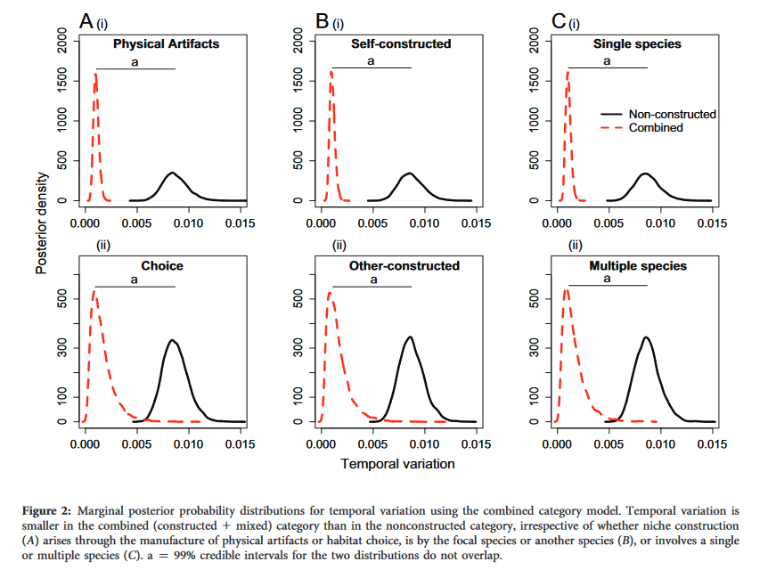
(2) Niche construction generates ecological inheritance
Ecological inheritance refers to the inherited resources and conditions, and associated modified selection pressures, that ancestral organisms pass on to their descendants as a direct or indirect result of their niche construction. For instance, if a beaver builds a dam transforming its local ecology, the modified selection will remain in the beaver’s environment just so long as the dam, lake, and modified environment persist, which can be decades. Likewise, the changes that earthworms produce in the soil can last many generations and can increase the fitness of the descendants (see Blog: Evolution’s Engineers).
It is well-recognized that environments can exhibit constancy across generations, but this is not generally viewed as an inheritance. Yet these ecological legacies have been shown to affect evolutionary dynamics strongly, and to contribute to parent-offspring similarity (Odling-Smee et al. 2003; Badyaev & Uller 2009; Odling-Smee et al. 2013). The stable inheritance of traits results in part from parents constructing developmental environments for their offspring (Badyaev & Uller 2009).
In recent years many evolutionary biologists have sought to expand the concept of inheritance within evolutionary biology, and ecological inheritance is now commonly incorporated into these schemes (Danchin et al. 2011; Bonduriansky 2012).
(3) Learning can be evolutionarily significant
There is considerable interest among evolutionary biologists in the role that imprinting, song learning, habitat imprinting, cultural transmission and various other forms of learning, play in speciation, the evolution of adaptive specializations, adaptive radiations, the colonization of new habitats, brood parasitism and sexual selection in vertebrates (ten Cate 2000; Laland et al. 2019; Whitehead et al. 2019).
From the niche-construction perspective, acquired characters, such as learned behaviour, can be evolutionarily important. Social learning in particular, is likely to exert a widespread influence on animal evolution. For instance, different clans of killer whales feed on very different prey species, with individuals learning their dietary preferences from older group members. As a consequence, clans have evolved specific jaws and digestive systems adapted to cope with their learned diets. Killer whales may currently be evolving into multiple separate species because of their cultural differences (Foote et al. 2016).
The learning of one species can influence the evolution of another. Reed warblers, for instance, learn to recognize cuckoos as brood parasites by attending to the alarm calls of other birds, a knock-on consequence of which is that natural selection favors cuckoos with unusual plumage patterns (Thorogood & Davies 2012). This gene-culture coevolution is of particular relevance to human evolution (Laland et al. 2010).

(4) Byproducts are evolutionarily significant
The niche-construction perspective highlights the important roles that byproducts can play in ecosystems. Such roles are not intuitive. For instance, it is far more apparent that the beaver’s dam may drive coevolutionary episodes than beaver’s dung may, yet the latter is a very real possibility. Numerous examples have been documented of seemingly inconsequential and inadvertent acts by organisms whose aggregate activity generates important consequences. For example, consider Euchondrus snails whose consumption of endolithic lichens inadvertently generates tonnes of soil, thereby playing a vital role in desert ecosystems (Jones & Shachak 1990).
Typically, biologists assume that if a niche-constructing activity generates evolutionary feedback to the constructor, then it must be an adaptation. In fact, theory shows this need not be the case. Byproducts can induce selection on other traits in the same population and hitchhike to fixation on the back of this selection (Silver & Di Paolo 2006). Here, spatial structure (local dispersal and mating) gives rise to statistical associations between niche-constructing traits and genotypes favored in the constructed environments. There is selection of the niche-constructing trait, but not selection for it, and only the latter meets the definition of an adaptation (Williams 1966; Sober 1984). Nonetheless, in such hitchhiking cases, there remains evolutionarily consequential feedback to a niche-constructing population stemming from its constructing activities.
(5) Niche construction creates a second route to adaptation
Evolutionary theory has historically focused on how organisms are shaped by natural selection to become suited to their environments. The niche-construction perspective emphasizes that through niche construction, environments can be changed by organisms to suit themselves. For instance, Turner (2000) notes that, despite living on land for millions of years, earthworms have retained the physiology characteristic of the freshwater worms from which they evolved. The earthworms process the soil in ways that allow them to draw water into their bodies more effectively, constructing a simulated aquatic environment on land. The adaptive complementarity of earthworms and soils results to a large extent from the worms changing the soil through niche construction, rather than the worms evolving a typical terrestrial physiology through natural selection.
These findings have led to the claim that niche construction is more than just a product of evolution, or source of environmental change, but should be recognized as a causal evolutionary process through its guiding influence on selection.
(6) Niche construction opens up new ecological niches
Through their niche construction, organisms can open up new ecological niches, both for themselves and for other species. Experiments have shown that niche construction evolves rapidly, under a broad range of conditions (Callahan et al. 2014), often leading to the creation of new niches. For instance, San Roman & Wagner’s (2018) experimental evolution investigation in bacteria showed that huge biodiversity could emerge in a completely homogeneous environment through niche construction. They found that bacteria created new ecological niches when they excrete nutrient-rich waste products that could sustain other bacteria, and identified thousands of such niches that had been created in this manner. Rather than lineages simply diversifying to “fill” available niches, niches themselves may be diversifying (Erwin 2005). Recent theoretical work shows that the construction of new niches by organisms has long-term, macro-evolutionary effects, for instance, increasing the branching patterns of phylogenetic trees (Xue et al. 2020).

 Blog article: “Is social transmission a forgotten force in coevolution?” by Rose Thorogood
Blog article: “Is social transmission a forgotten force in coevolution?” by Rose Thorogood


Researchers studying niche construction commonly embrace an alternative perspective in which niche construction is regarded as a fundamental evolutionary process in its own right. Evolution entails networks of causation and feedback in which previously selected organisms drive environmental changes, and organism-modified environments subsequently select for changes in organisms.
The perspective is intellectually aligned with several other movements within evolutionary biology, including ‘developmental systems theory’ (Oyama et al. 2001), the active role of behavior (Bateson 1988), and of developmental plasticity (West-Eberhard 2003), in evolution, and calls for an extended evolutionary synthesisA new evolutionary framework emphasizing that knowledge of how organisms develop, grow, and interact with environments helps to account for adaptation and the diversity of life. (Pigliucci & Muller 2010; Laland et al. 2015).
The extended evolutionary synthesis, or EES, is a new way to think about and understand evolutionary phenomena that differs from the conception that has dominated evolutionary thinking since the 1930s (i.e. the modern synthesis). It is a developing line of contemporary evolutionary thought that exists within the field of evolutionary biology.

According to the EES, explaining the origin of adaptations requires understanding how developmental processes generate heritable phenotypic variants from genetic, epigenetic and environmental inputs. Developmental biasThe nonrandom generation of phenotypes by developmental systems, with variants sometimes being channeled by the processes of development towards functional goals. and phenotypic plasticityEnvironmental induction leads to developmental reorganization and production of a novel phenotypic variant. play central roles in the EES as generators of novel, yet potentially functional and coordinated, phenotypic variation. This conception of bias is different from the traditional characterization of developmental constraints: rather than accounting for the absence of evolution or adaptation, developmental bias is also a source of adaptive variation.
Developmental bias and niche construction are, in turn, recognized as evolutionary processes that can initiate and impose direction on selection. Extra-genetic inheritanceParental transference of developmental resources (mediated through genetic, epigenetic, physiological, behavioural and ecological inheritance mechanisms) that enable reconstruction of life cycles. mechanisms (which include ecological inheritance) interact with genetics and environmental inputs to construct the developing organism, thereby contributing to the similarity between ‘transmitting’ and affected individuals.
Like niche construction theory, the EES emphasizes how organisms shape and are shaped by selective and developmental environments (reciprocal causationProcess A is a cause of process B and, subsequently, process B is a cause of process A. Reciprocal causation captures the idea that developing organisms are not solely products, but are also causes, of evolution.), and how the development of organisms is not programmed but open-ended and constructive (constructive developmentThe developing organism shapes its own developmental trajectory by responding to environmental inputs and altering internal and external states.). Developmental processes, operating through developmental bias and niche construction, are viewed as sharing with natural selection some responsibility for the direction and rate of evolution and contribute to organism–environment complementarity.
The resulting network of processes provides a considerably more complex account of evolutionary mechanisms than traditionally recognized. The EES entails not only new research directions but also new ways to think about, and interpret, new and familiar problems in evolutionary biology.
Check out the Extended Evolutionary Synthesis resource website:
![]()