Domestication and Agriculture are the Outcome of Plant Gene-Human Culture Coevolution
11 January 2020
In our recent paper, “Understanding agriculture within the frameworks of cumulative cultural evolution, gene-culture coevolution and cultural niche construction” (Human Ecology 47:483–497), we apply the framework and concepts of Gene-Culture Coevolution (GCC), Niche Construction (NC) and Cumulative Cultural Evolution (CCE) to agriculture. All three concepts were originally conceived with respect to human culture and human genes, and later on also with animal genes (mainly domesticated animals). However, to the best of our knowledge, ours is the first full discussion of how plant genes coevolve with human cultural practice.
Beginning around 12,000 years ago, some human populations began domesticating plant and animal species1-2. The transition from hunting and gathering to agriculture is a key event in human history, and the adoption of agriculture is believed to have triggered the establishment of small permanent settlements, and, as populations expanded, the evolution of cities, kingdoms, and states. Agriculture stimulated the creation of new political institutions and forms of social organization, and an upsurge in scientific and technological innovation.
One key result was improved food production and the supply of more food to more people. Agricultural knowledge and technologies have continued to advance at an increasing pace, particularly in the last century. The discovery of the rules of genetics by Mendel3 and their rediscovery around 1900 resulted in the application of plant breeding technologies from the 1930s onwards. The “green revolution” in several developing countries during 1950–1970, which utilized new high yield crops together with fertilizers and pesticides, was an important landmark in agricultural plant breeding, yet was still based on traditional Mendelian breeding methods4. The more effective new technologies resulted in higher crop yields, yet populations increased at a higher pace. The demand for food often surpassed crop production, especially in developing countries. The era of molecular breeding, including marker-assisted selection (MAS) brought about new possibilities, and was followed from 1983 onwards by widespread genetic engineering/modification of crop plants5, and more recently by genome editing technologies6-7. Molecular breeding has transformed agricultural practices worldwide, although it often faces strong public and political opposition.
Despite the importance of agriculture to our species’ history and recent rapid advances in molecular breeding technologies, there remain disagreements over which theoretical framework offers the best understanding of the origin, spread, and ongoing transformation of agriculture. Several recent debates and exchanges have revealed a tension between, on the one hand, interpretive, humanities-oriented frameworks that focus on culture and agency on the part of agriculturalists and the sociopolitical contexts within which agriculture is practiced and, on the other hand, neo-Darwinian approaches that use tools such as optimal foraging theory derived from behavioral ecology to understand agricultural decisions, assuming that human decision-making has genetically evolved to maximize inclusive genetic fitness.
Here we follow others8–10 in arguing that the study of agriculture can benefit from being situated within a set of new evolutionary approaches to human behavior – cultural evolution, gene-culture co-evolution, and cultural niche construction – that attempt to incorporate cultural change and individual agency within a rigorous scientific and multidisciplinary evolutionary framework. We point out that changes in agricultural knowledge and practices are a prime example of cumulative cultural evolution (CCE) and gene-culture co-evolution (GCC), and that agriculture is associated with extensive cultural niche construction (CNC), in part due to the fact that agricultural practices transform the environment and those environmental changes alter the selection pressures on agricultural CCE.
For example, agriculture can modify the abiotic environment (e.g., water, salinity, soil composition), the biotic environment (e.g., domesticated species, pests including insects, fungi, and weeds), and the social environment (e.g., social norms, regulation, markets). As such, agriculture is one of the major causes of the Anthropocene. Moreover, the recognition that domestication, breeding, and the advancement of agriculture are the outcome of plant gene-human culture coevolution are most relevant to the many issues of public acceptance and spread of the new genetically modified crop plants.
Most previous discussion of GCC in the context of agriculture concerns deep human history and prehistory (e.g., lactose tolerance and dairy farming; O’Brien and Laland 2012). In this paper, we instead focus on recent biotechnology and molecular breeding to illustrate the points raised above and demonstrate the relevance of these theoretical frameworks to contemporary issues. The molecular breeding technology described above is clearly a case of CCE, building on what went before (e.g., Mendelian genetics) and far exceeding what any single individual could achieve alone. This exponential accumulation of knowledge is a well-recognized characteristic of CCE11.
Molecular breeding is also a case of GCC, where the genes of other species (e.g., plants) are directly and intentionally modified using culturally evolving scientific techniques, unlike (and much faster than) undirected natural selection and traditional artificial selection. These genetic modifications in turn demand new and more powerful scientific techniques and knowledge.
Finally, molecular breeding involves extensive CNC, in terms of major changes to the abiotic, biotic, and social environments. This was analyzed in our publication by carefully examining relevant practical characteristics in three outstanding case studies of genetically-modified (GM) crop plants: increased vitamin A production in “golden rice”, and the generation of herbicide and insect tolerant crops. We described both the needs for engineering for the mentioned plant characteristics and the relevant technologies/scientific procedures involved, and analyzed the evidence for the variable associated involvement of socio-cultural factors (including, for example, public trust, ethical issues, different culinary habits, gender issues and more), as well as cross-cultural differences in acceptance or rejection of GM foods.
All various agricultural technologies, ancient and novel, are clearly the result of CCE, with each major advance dependent on previous advances. Molecular breeding and genetic engineering could not have been invented without existing knowledge of Mendelian genetics. Yet, there are differences. The Neolithic agricultural period, i.e., plant and animal domestication, as well as other technological improvements in agriculture and biology (e.g., the use of irrigation and fertilizers) are more protracted and evolved sequentially over a period of hundreds or thousands of years. In contrast, the time span of adopting and applying molecular plant breeding technologies and in vitro biology has been much shorter. Such technologies emerged far more rapidly, and became a working reality only within the last few decades. The molecular structure of DNA was first published in 195312, and the first genetically modified (GM) or transgenic plant (i.e., produced via incorporation of recombinant DNA), tobacco, was first created in the laboratory in 1982 5, 13-15. Farmers began to plant GM crops in 1996, and in 2017, the 21st year of commercialization of biotech crops, 189.8 million hectares (a ~112-fold increase) of biotech crops were planted by up to 17 million farmers in 24 countries, which makes GM crops the fastest adopted crop technology in recent times16-21.
All three case studies of genetically-modified crops that we studied, and many other GM plants, indicate that the evolution of agricultural practices are inseparable from human cultural evolution, are associated with human and environmental niche construction, and are the result of the overall gene culture coevolution that includes human, animal and plant genes.
First, we argue that agriculture is an excellent case of CCE. It involves the sequential improvement over time of agricultural knowledge (both scientific and non-formal knowledge systems) and practices (from small-scale habits and routines to large-scale technology) via the repeated cycle of innovation and cultural transmission. Viewing changes in agricultural practices as an evolutionary process and recognizing the resultant co-evolutionary dynamics and feedbacks facilitates connecting this cultural process with the biological/evolutionary/natural sciences, preventing a false and unproductive nature-culture dichotomy. Agriculture informally exhibits the classic exponential increase in knowledge and practices that is typical of CCE, with recent change seemingly orders of magnitude faster than past rates of change, allowing the large body of work exploring the drivers and inhibitors of CCE to further contribute to agricultural research.
Second, we argue that the standard notion of GCC, where human cultural practices shape human genes and vice versa should be expanded to include culturally-driven changes in non-human genes. This includes, by definition, domestication, which entails the traditional breeding of domesticated species. More recently this has involved direct genetic modification with the introduction of GM crops.
Third, agriculture is a prime example of CNC, involving extensive modification of abiotic, biotic, and social environments, and feedback from these environments to agricultural knowledge and practices. Most interesting from our perspective are feedbacks with the social environment. Adoption of golden rice and other GM crops has generated resistance from activist groups, political parties, and regulators due to fears over food safety, genetic contamination, and an aversion to ‘tampering with nature.’ These concerns provoke increased regulation and safety testing within the agriculture industry to ensure that GM products are as safe as possible. While adequate levels of health regulation are of course needed, overly stringent regulation can prevent potentially beneficial innovations from spreading. The ideal outcome would be improved population health and reduced environmental impact as a result of GM crops such as golden rice, green super rice, and herbicide/insect resistant plants, as well as drought and salinity tolerant crops, post-harvest loss of food, use of novel fertility control in farm animals and more. Another positive social feedback is the impact on gender roles, with herbicide tolerant GM crops releasing women in many low-income countries from excessive manual labour (e.g. weeding) and improving educational and economic outcomes. Thus, while gene-culture coevolution is typically conceived as culturally-driven change in human genes, we extend it to include also genetic changes in domesticated plants and animals.
This is further illustrated schematically in the following figure:
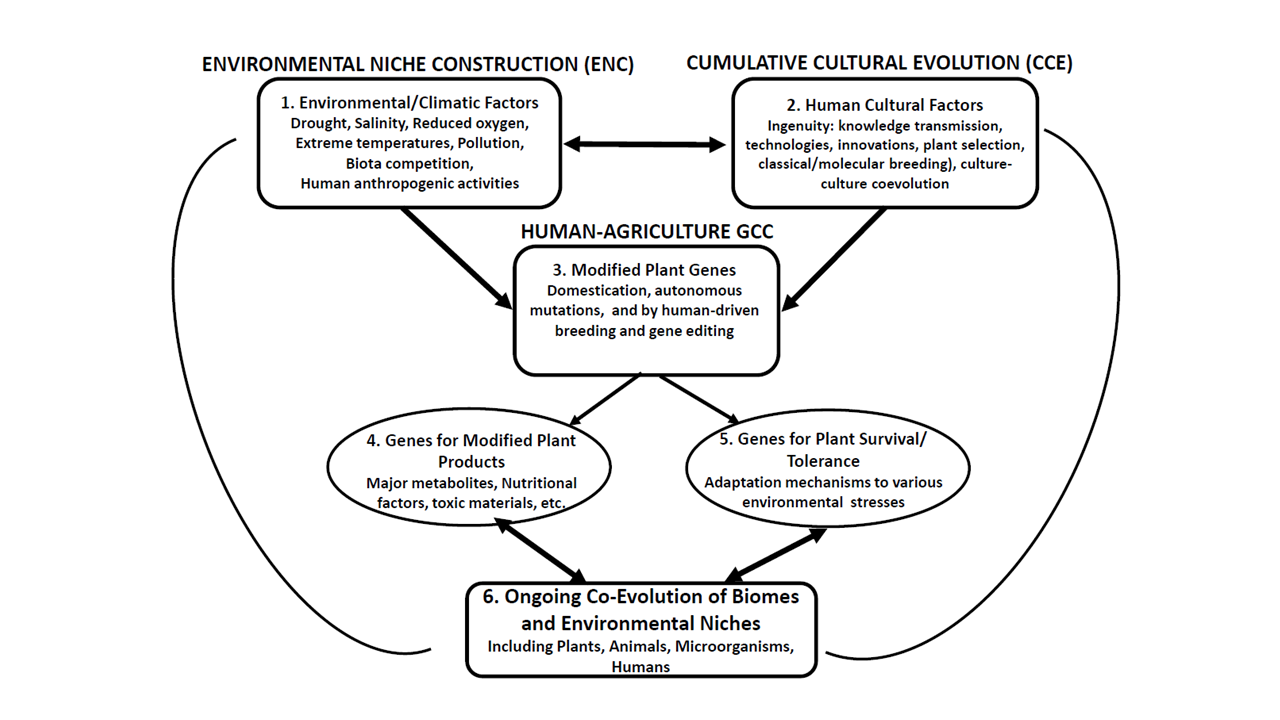
Figure. Major agriculture and culture-associated niche construction and plant gene-culture coevolution. The different interacting components of cumulative cultural evolution (CCE), plant-specific gene-culture coevolution (GCC), and environmental/agriculture associated cultural niche construction (CNC) are schematically represented. Two major components are implicated: the physical environment, i.e., geography, the terrain, climate, and more (Box 1), human cultural factors, including ingenuity, technology and scientific discoveries (Box 2). Both may modify, shape, interact and coevolve with specific genes of domesticated plants (and farm animals) (Box 3). Once a certain selected gene combination has been fixed in a domesticated plant (or a farm animal) it can be again modified by traditional breeding techniques or by employment of novel molecular tools (MAS, GM, Genome editing) to produce novel gene combinations affecting mainly genes associated with modified plant products and metabolites (Box 4) and genes for improving plant survival/ tolerance to environmental stresses (Box 5). The novel plant products or traits can in turn result in the creation of new environmental niches, affect the expression of human genes through consuming those products, resulting in ongoing coevolution of biomes (i.e., the entire complex body of living organisms including plants, animals, and microorganisms), CCE, GCC, and ENC (Box 6)
To conclude, early plant domestication and transition to agriculture is a key event in human history, followed by traditional breeding, and currently also by genetic engineering and gene editing – all dependent on the invention and diffusion of novel technologies via a process of cumulative cultural evolution. We analyze several case studies of genetically-modified plants and discuss evidence for cross-cultural differences in acceptance or rejection of GM foods. We argue that agriculture can benefit from a set of new evolutionary approaches to human behavior, suggesting that changing agricultural practices are a prime example of cumulative cultural evolution (CCE), are associated with specific niche construction (NC), and represent a specific case of gene-culture coevolution (GCC). While gene-culture coevolution is typically conceived as culturally-driven change in human genes, we extend it to include also genetic changes in domesticated plants and animals. Overall, we propose new insights into the co-evolution of human culture, plant genes and agriculture and plant genes and the unprecedented contribution of agricultural activities at large that had transformed our species and our planet to such an extent that it is one of the primary attributes of the Anthropocene.
******
This publication is the outcome of true collaborative research between experts from the two disciplines, the “gene” and the “culture”. Arie Altman in agriculture and plant biology and biotechnology, and Alex Mesoudi in gene-culture coevolution and cumulative cultural evolution. Together we examined several case studies of GM plants, related to the fact that both plant domestication and, especially, continuous plant breeding (including recent molecular breeding) are the result of human activities. Moreover, in this publication we expand the original scope of the term Gene-Culture Coevolution and propose its relevance to many other genes, including plant genes, rather that only human, i.e., we should thus refer to Plant Gene-Human Culture Coevolution. This new concept fits well within the extended evolutionary synthesis framework.
For more details, read the paper here:
Altman A, Mesoudi A. 2019. Understanding Agriculture within the Frameworks of Cumulative Cultural Evolution, Gene-Culture Co-Evolution, and Cultural Niche Construction. Hum Ecol. 47(4):483–97. [download pdf]
Arie Altman (Professor of horticulture and agricultural biotechnology at the Faculty of Agriculture, Food and Environment of the Hebrew University of Jerusalem, Israel and Invited Professor at the Culture Research Unit , Faculty of Humanity of Tel Aviv University, Israel) has recently written another paper on plant gene-human culture coevolution, together with Stephen Shennan (UCL, London, UK) and John Odling-Smee (Oxford University, UK) entitled “Domestication and gene-culture coevolution of ornamental plants as a specific aesthetics-driven component of the human niche”. In this manuscript we examine the socio cultural reasons for domestication and breeding of ornamental plants, that unlike food plants were not domesticated for “physical” needs like food. We studied several key ornamental plant species and conclude that their domestication and breeding, however, is a specific aesthetics-driven dimension of human niche construction that has coevolved with socio-economic changes, new scientific technologies and symbolic-material asset shifts, with a foundation in human sense of beauty and aesthetic values to which ornamental plants appeal. We further argue that archaeological records and the cultural histories of ornamental plants indicate that people started to cultivated them about 4,000 years later than food plant domestication, and that this event was probably associated with the rise of elites whose power and wealth were used to support craft-specialization for the production of new highly elaborate forms of material culture. Within the plant biotechnology field of study, he and colleagues published recently on Accelerating Climate Resilient Plant Breeding by Applying Next-Generation Artificial Intelligence. 2019. Trends in Biotechnology 37: 1217-1235. He is preparing currently another publication on “Promoting Ethically and Socially Responsible Use of Plant Biotechnology: Towards Public Acceptance”.
Alex Mesoudi (Professor of Cultural Evolution, University of Exeter, UK) currently has several projects empirically and theoretically examining different aspects of human cultural evolution. One ongoing project is examining immigrant acculturation through the lens of cultural evolution theory. This involves modelling acculturation, i.e. the changes in psychological and behavioural characteristics that result from migration, in terms of vertical, oblique and horizontal cultural transmission pathways (Mesoudi 2018), and empirically studying acculturation pathways in British Bangladeshis in London (Mesoudi, Magid & Hussain, 2016). Another project is experimentally simulating the process of ‘prestige bias’, where people preferentially learn from high status individuals (Brand & Mesoudi 2019; Jimenez & Mesoudi 2019). Other experimental work has examined the dynamics of cumulative cultural evolution in the lab, in particular whether it requires explicit understanding of how technology works (Derex, Bonnefon, Boyd & Mesoudi 2019).
Further readings:
- Boivin, N. L., Zeder, M. A., Fuller, D. Q., Crowther, A., Larson, G., Erlandson, J. M., and Petraglia, M. D. (2016). Ecological Consequences of Human Niche Construction: Examining Long-Term Anthropogenic Shaping of Global Species Distributions. Proceedings of the National Academy of Sciences 113(23): 6388–6396.
- Bonny, S. (2016). Genetically Modified Herbicide-Tolerant Crops, Weeds, and Herbicides: Overview and Impact. Environmental Management 57(1): 31–48.
- Cavalli-Sforza, L. L., and Feldman, M.W. (1981). Cultural Transmission and Evolution, Princeton Univ. Press, Princeton.
- Echols, M. A. (1998). Food safety Regulation in the European Union and the United States: Different Cultures, Different Laws. Columbia Journal of European Law 4: 525–543.
- Ellis, E. C., Magliocca, N. R., Stevens, C. J., and Fuller, D. Q. (2018). Evolving the Anthropocene: Linking Multi-Level Selection with Long-Term Social-Ecological Change. Sustainability Science 13: 119–128.
- Enquist, M., Ghirlanda, S., and Eriksson, K. (2011). Modelling the Evolution and Diversity of Cumulative Culture. Philosophical Transactions of the Royal Society B 366: 412–423.
- Farre, G., Ramessar, K., Twyman, R. M., Capell, T., and Christou, P. (2010). The Humanitarian Impact of Plant Biotechnology: Recent Breakthroughs vs Bottlenecks for Adoption. Current Opinion in Plant Biology 13: 219–225.
- Feldman,M.W., and Laland, K. N. (1996). Gene-Culture Coevolutionary Theory. Trends in Ecology and Evolution 11(11): 453–457.
- Fuller, D. K., Willcox, G., and Allaby, R. G. (2012). Early Agricultural Pathways: Moving Outside the ‘Core Area’ Hypothesis. Journal of Experimental Botany 63: 617–633.
- Gasser, C. S., and Fraley, R. T. (1989). Genetically Engineering Plants for Crop Improvement. Science 244(4910): 1293–1299.
- Heslop-Harrison, J.S. and Scwarzacher, T. (2012). Genetics and genomics of crop domestication. In: A. Altman and P.M. Hasegawa eds., Plant Biotechnology and Agriculture: Prospects for the 21st Century. London: Elsevier/Academic Press. ISBN-10: 0123814669. pp. 3-18.
- ISAAA (2017). Global Status of Commercialized Biotech/GM Crops in 2017: Biotech crop Adoption Surges as Economic Benefits Accumulate in 22 Years. ISAAA Brief No. 53. ISAAA: Ithaca, NY.
- Laland, K. N., Odling-Smee, J., and Myles, S. (2010). How Culture Shaped the Human Genome: Bringing Genetics and the Human Sciences Together. Nature Reviews Genetics 11(2): 137–148.
- Odling Smee, F. J., Laland, K. N., and Feldman, M. (2003). Niche Construction, Princeton University Press, Princeton.
- Potrykus, I. (2010). Lessons from the ‘Humanitarian Golden Rice’ Project: Regulation Prevents Development of Public Good Genetically Engineered Crop Products. New Biotechnology 27: 466–472.
- Richerson, P. J., Boyd, R., and Henrich, J. (2010). Gene-Culture Coevolution in the Age of Genomics. Proceedings of the National Academy of Sciences 107(Supplement 2): 8985–8992. https://doi. org/10.1073/pnas.0914631107.
- Rowley-Conwy, P., and Layton, R. (2011). Foraging and Farming as Niche Construction: Stable and Unstable Adaptations. Philosophical Transactions of the Royal Society B: Biological Sciences 366(1566): 849–862.
- Sander, J. D., and Joung, J. K. (2014). CRISPR-Cas Systems for Editing, Regulating and Targeting Genomes. Nature Biotechnology 32: 347–355.
- Slingerland, E., and Collard,M. (2011). Creating Consilience: Integrating the Sciences and the Humanities, Oxford University Press.
- Smith, B. D. (2011). General Patterns of Niche Construction and the Management of ‘Wild’plant and Animal Resources by Small-Scale Pre-Industrial Societies. Philosophical Transactions of the Royal Society of London B: Biological Sciences 366(1566): 836–848.
- Subramanian, A., Kirwan, K., Pink, D., and Qaim,M. (2010). GMCrops and Gender Issues. Nature Biotechnology 28: 404–406.
- Zeder, M. A. (2015). Core Questions in Domestication Research. Proceedings of the National Academy of Sciences 112(11): 3191–3198. https://doi.org/10.1073/pnas.1501711112.
1. Fuller DQ, Denham T, Arroyo-Kalin M, Lucas L, Stevens CJ, et al. 2014. Proc. Natl. Acad. Sci. 111(17):6147–52. 2. Larson G, Piperno DR, Allaby RG, Purugganan MD, Andersson L, et al. 2014. Proc. Natl. Acad. Sci. 111(17):6139–46. 3. Mendel GJ. 1866. Verhand-Lungen Naturforschenden Vereines Brünn Bd IV Für Jahr 1865. IV (1865):3–47. 4. Farmer BH. 1986. Mod. Asian Stud. 20(1):175–99. 5. Gasser CS, Fraley RT. 1989. Science. 244(4910):1293–99. 6. Sander JD, Joung JK. 2014. Nat. Biotechnol. 32(4):347. 7. Bortesi L, Fischer R. 2015. Biotechnol. Adv. 33(1):41–52. 8. Rowley-Conwy P, Layton R. 2011. Philos. Trans. R. Soc. B Biol. Sci. 366(1566):849–62. 9. O’Brien MJ, Laland KN. 2012. Curr. Anthropol. 53(4):434–70. 10. Zeder MA. 2015. Proc. Natl. Acad. Sci. 112(11):3191–98. 11. Enquist M, Ghirlanda S, Jarrick A, Wachtmeister C-A. 2008. Theor. Popul. Biol. 74(1):46–55. 12. Watson JD, Crick FHC. 1953. Nature. 171(4356):737–38. 13. de Framond AJ, Barton KA, Chilton M-D. 1983. Bio/technology. 1(3):262. 14. Zambryski P, Joos H, Genetello C, Leemans J, Van Montagu M, Schell J. 1983. EMBO J. 2(12):2143–50. 15. Tepfer D. 1984. Cell. 37(3):959–67. 16. Farre G, Ramessar K, Twyman RM, Capell T, Christou P. 2010. Curr. Opin. Plant Biol. 13(2):219–25. 17. Altman A, Hasegawa PM. 2012. Plant Biotechnology and Agriculture: Prospects for the 21st Century. Academic Press. 18. Altman A, Hasegawa PM. 2012. In Plant Biotechnology and Agriculture, ed A Altman, PM Hasegawa. xxix–xxxviii. San Diego: Academic Press. 19. Moshelion M, Altman A. 2015. Trends Biotechnol. 33(6):337–42. 20. ISAAA. 2017. In ISAAA Brief No. 53. Ithaca, NY.: ISAAA. 21. Harfouche AL, Jacobson DA, Kainer D, Romero JC, Harfouche AH, et al. 2019. Trends Biotechnol.